LUYOR-3109高強度紫外催化光源促銷
LUYOR-3109紫外光源采用了9顆365nm大功率led,安裝有二次光學透鏡,輸出紫外線強度高,...
2024-08-08

作者:激發光源事業部時間:2019-12-08 11:32:28瀏覽4285 次
天然gfp的熒光強度低,表達易受溫度影響,通過改變發色團氨基酸種類和排列順序,獲得了改良型的gfp。將第65位的ser突變成thr,可顯著增加熒光強度和穩定性;第66位氨基酸tyr突變為his或trp,熒光波長變短,可分別得到發藍光和藍綠光的gfp;改變發色團周邊氨基酸的種類,也可以使熒光波長發生變化,將第203位的thr突變成tyr,就可獲得黃色熒光。
綠色熒光蛋白在植物病理學研究中的應用
報告基因(reportergene)是編碼某種檢測蛋白或酶的基因,通過它的表達產物來標定目的基因的表達調控情況,具有靈敏度高、可信度好和檢測方便。報告基因種類繁多,如氯霉素乙酰轉移酶(chloramphenicolacetyltransferase,cat)基因、分泌型堿性磷酸酶(secretedalkalinephosphatase,seap)基因、綠色熒光蛋白(green fluorescentprotein,gfp)基因、β-葡萄糖苷酶(β-glucosidase,gus)基因、β-半乳糖苷酶(β-galactosidase,β-gal)基因和螢火蟲熒光素酶(fireflyluciferase,fl)基因等。由于gfp具有熒光性質穩定、觀察方便、對細胞無毒害、無物種特異性、不受假陽性干擾和無需底
物等優點,深受科研和檢測人員的青睞,目前已成為分子生物學領域的重要標記蛋白。
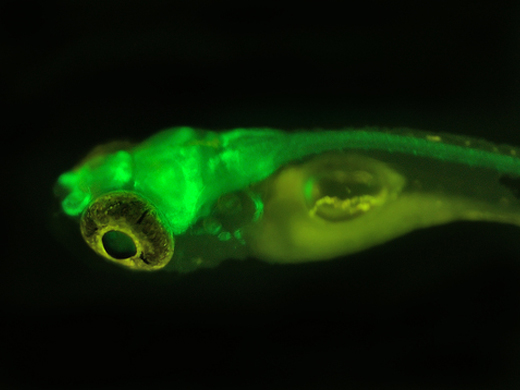
綠色熒光蛋白于1962年在維多利亞多管發光水母(aequoreavictoria)中發現,該蛋白在紫外光照射下發出明亮的綠光;1992年prasher成功克隆gfp的全長cdna,2年后將gfp基因作為報告基因導入大腸桿菌(escherichiacoli)和秀麗隱桿線蟲(caenorhabditiselegans)中表達,發現在無任何輔助底物的情況下,gfp可以在活細胞中發光;通過基因突變和化學修飾對gfp基因進行改造,解決了gfp熒光強度低、光選擇性差和光譜狹窄等缺陷,目前熒光顏色已涵蓋紫色到紅色的所有可見光區域。近年來,gfp基因在啟動子分析、轉基因監測、藥物檢定、污染物示蹤、抗病檢測和基因表達調控等方面得到了廣泛應用。gfp基因在植物病理學研究中也得到了廣泛應用,本文將展開相關論述。

gfp的發現
生物發光是一種普遍現象,自然界中,大約有700種生物能夠發光,包括細菌、大型真菌和動物,約80%的發光生物生活在海洋中,它們通過發光來尋找食物、逃避敵害和吸引異性。生物發光通常需要底物的參與,如螢火蟲發光是通過熒光酶)催化熒光素(luciferin)來實現的,而gfp是一種能在藍色光線激發下發出綠色熒光的特殊蛋白質,在發現初期并未受到重視,它只是研究水母素(aequorin)時的副產品。20世紀90年代,gfp在大腸桿菌中成功表達,開創了該蛋白的應用先河,之后在生命科學多個領域中得到了廣泛應用,并取得了令人矚目的成就。
gfp的結構與發光原理
野生型gfp基因組全長為2600bp,由3個外顯子和2個內含子組成,編碼區全長為717bp,分別以atg和taa為起始和終止密碼子,編碼238個氨基酸,分子量為27ku。gfp的三維結構呈圓筒形,11個β折疊圍在四周,1個α螺旋貫穿整個圓筒結構,其中3個氨基酸殘基ser65-tyr66-gly67構成發色團,位于圓筒中央并附著在α螺旋上[12]。盡管在氨基酸水平上僅23%的相似性,但gfp的結構與從discosomasp.分離到的紅色熒光蛋白(redfluo-rescentprotein,rfp)十分相似,后者也是由11個β折疊和1個α螺旋組成,形狀也呈折疊筒狀[12]。gfp的發光原理是:gly67的酰基和ser65的羧基經親核反應生成咪唑基,tyr66通過脫氫使芳香團與咪唑基結合,形成對羥基苯甲酸咪唑環酮生色團發出熒光。
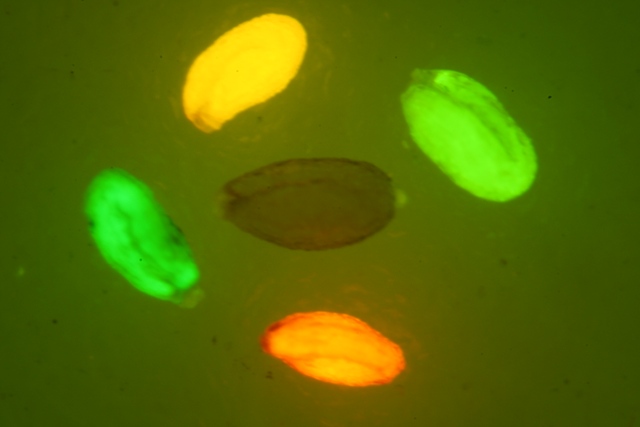
天然gfp的熒光強度低,表達易受溫度影響,通過改變發色團氨基酸種類和排列順序,獲得了改良型的gfp。將第65位的ser突變成thr,可顯著增加熒光強度和穩定性;第66位氨基酸tyr突變為his或trp,熒光波長變短,可分別得到發藍光和藍綠光的gfp;改變發色團周邊氨基酸的種類,也可以使熒光波長發生變化,將第203位的thr突變成tyr,就可獲得黃色熒光。
gfp在植物病理學研究中的應用
隨著成像技術、檢測手段的快速發展,gfp基因及其突變體作為報告基因在分子生物學領域中得到了大規模應用。gfp基因在植物病理學方面主要體現在抗病基因的亞細胞定位、啟動子活性分析、病原菌-植物互作研究和基因表達分析等方面。
3.1抗病基因的亞細胞定位
真核細胞具有復雜亞細胞結構,蛋白質在細胞內的定位是細胞生物學研究的熱點之一。亞細胞定位能明確表達蛋白或產物在細胞內的出現部位,常用的方法有免疫熒光技術、膠體金標記免疫電鏡技術和gfp融合蛋白技術等。免疫熒光技術將免疫學方法與熒光標記技術相結合研究特異蛋白抗原在細胞內分布,通過檢測熒光素發出的熒光對組織或細胞中的抗原進行細胞定位,這種方法具有簡便、快速和特異等優點,缺點是敏感性低、定量困難和易受化學發光物質干擾[16]。膠體金標記免疫電鏡技術是以膠體金作為示蹤標志物或顯色劑,利用其在堿性條件下帶負電的特點進行抗體標記,具有特異性強、靈敏度高和定位正確等優點,缺點是實驗步驟多、技術難度大。而gfp具有熒光性質穩定、無物種特異性、對細胞無毒、靈敏度高和能在細胞中穩定表達等優點,gfp融合蛋白技術已成為亞細胞定位的重要手段。
npr1基因是擬南芥(arabidopsisthaliana)系統獲得抗性中的一個關鍵基因,它在植物抗病反應中起到重要作用,國內外許多學者對其進行了較系統的研究,利用農桿菌介導法將npr1-gfp融合基因導入擬南芥,結果表明,npr1基因在細胞核中表達,在病菌誘導下,npr1-gfp整合蛋白的積累量增加[17]。osWrky13是水稻(oryzasativa)的一個轉錄因子,過量表達能提高對白葉枯病和稻瘟病的抗性,將gfp-osWrky13融合基因導入洋蔥表皮,結果表明該基因在細胞核中表達,并進一步證明osWrky13具有dna結合活性[18]。過氧化物酶在抗病反應中起著重要的調控作用,將一粒小麥(triticummonococcum)過氧化物酶基因tmprX8與gfp融合后導入洋蔥表皮,融合蛋白出現在液泡中。
3.2啟動子活性分析
啟動子是一段直接與rna聚合酶及轉錄因子相結合,決定基因轉錄起始與否的特定dna序列,采用有效的啟動子能夠顯著提高外源基因在植物中的表達水平,因此啟動子活性檢測是植物基因工程的重要內容之一。傳統的啟動子活性檢測采用β-葡萄糖苷酸酶基因、氯霉素乙酰轉移酶基因和冠纓堿合成酶基因等作為報告基因,檢測時需要特定的底物和輔助因子,而且對植物組織或細胞具有破壞性。gfp是啟動子分析的新型報告基因,利用熒光顯微鏡觀察基因表達情況,能較正確地鑒定啟動子活性大小,并達到量化的目的。
蘇云金芽胞桿菌(Bacillusthuringiensis)是一種應用廣泛的殺蟲微生物,已成功地用于防治農業、林業、貯藏物害蟲及醫學昆蟲,隨著微生物農藥需求量的增大和人類環境保護意識的增強,生物安全日益受到人們的重視。利用gfp基因研究了蘇云金芽胞桿菌pcry3a和pBtⅠ_BtⅡ等啟動子的活性,為有效選擇驅動標記基因更優的表達體系和進一步構建、安全的基因工程菌創造了條件[21]。利用gfp基因作為報告基因,采用基因槍法進行洋蔥表皮遺傳轉化,培養24h后檢測基因表達情況和熒光強度,能正確測定出啟動子活性的大小。
3.3病原菌與植物互作研究
病原菌侵染植物過程中,病原菌無毒基因產物與寄主植物抗病基因產物相互識別和結合,產生的信號分子通過傳導,激活一系列抗病基因的表達,引發復雜的生理生化反應,從而表現出抗病特性[22]。深入研究植物與病原菌的識別方式、親和性或非親和性的互作模式,對揭示植物-病原菌互作機制的認識具有重要意義[23]。研究病原菌與植物互作的傳統方法有組織印跡法、放射性標記核酸探針法和gus染色法等,這些均為非活體檢測,因此有一定的局限性[21]。而gfp能有效克服這一弊端,可直觀、實時監測病原菌的發生、定殖和侵染過程,目前已成為研究病原菌與植物互作的有力工具[25]。煙草黑脛病是煙草生產過程中的重要病害,世界各地均有發生,利用原生質體介導法將gfp基因導入病原菌(phytophthoraimperfectavar.nicoti-anae)中,觀察到了煙草黑脛病菌的生長動態和與寄主互作方式,這是gfp作為報告基因在病原微生物研究中的應用。香蕉葉斑病和斑枯病由球腔菌屬(Mycosphaerella)真菌引起,其中香蕉黑條葉斑病菌(M.fijiensis)和香蕉黃條葉斑病菌(M.musi-cola)為葉斑病的病原菌,而M.eumusae為斑枯病的病原菌,將gfp基因導入這3種真菌,清楚地觀察到了病菌在葉片組織中的生長動態,并且發現隨著菌絲的侵入葉片逐漸褪綠和壞死。草莓炭疽病是草莓(fragariaananassa)生產的重要病害,近年來發生十分嚴重,尖孢炭疽菌(colletotrichumacutatum)是草莓炭疽病的病原菌,它侵染葉片、匍匐莖、花蕾、萼片和果實,用電穿孔法將gfp基因導入萌發的分生孢子研究草莓致病炭疽病原菌和非致病炭疽病病原菌的生活規律,為闡明致病機理奠定了基礎。
利用含gfp基因和氯霉素抗性的質粒分別轉化對黃瓜枯萎病菌(fusariumoxysporum)有明顯抑制作用的生防菌株BrevibacillusbeviszJy1和BacillussubtiliszJy-116,得到具有gfp綠色熒光和氯霉素抗性的轉化子,成功檢測了菌株在田間定殖的可行性,為菌株的實際開發利用提供了依據。以萵苣盤梗霉(Bremialactucae)hsp70的啟動子和ham34基因的終止子構建gfp表達載體,利用原生質體介導法導入重組載體,觀察到gfp在不同時期大豆疫霉(phytophthorasojae)中的表達情況,同時明確了大豆疫霉在大豆葉片、下胚軸和根部的侵染行為新霉素磷酸轉移酶基因(hpt)、潮霉素磷酸轉移酶基因(npt)、鏈霉素磷酸轉移酶基因(nptii)、草銨膦(glufosinate)抗性基因(bar)、草甘膦(glyphosate)抗性基因(epsps)等為傳統的轉基因植株選擇標記[32],通常需要6~8周或更長的時間才能鑒定出潛在的轉基因苗,并經southern雜交分析來確定是否為轉基因陽性植株,整個過程花費時間較長。使用抗生素、除草基因作為轉基因標記是否會引發生物安全問題是人們普遍關注的問題,而gfp基因是一種安全、可靠、省時的標記基因,不會產生“超級雜草”和“基因逃逸”等生態問題,可替代抗素、除草劑快速篩選轉基因植株或組織,還可通過標記花粉來研究轉基因植物花粉漂移和授粉過程。
sgt1(suppressoroftheg2alleleofskp1)是植物抗病信號傳導途徑中的重要調控基因,參與多種植物抗病基因介導的抗病信號途徑,sgt1基因的突變或沉默會導致多種植物r基因介導的抗病性的喪失,sgt1還參與調控植物的非宿主抗性[34-35]。以gfp為報告基因對atsgt1a和ats-gt1b的功能進行了研究,通過熒光檢測,明確了這兩個抗病基因的表達部位和表達量[36]。利用peg法將來自水稻的抗病基因Xa21導入柑橘原生質體,通過gfp熒光檢測獲得愈傷組織和抗性芽,經微嫁接后獲得了抗病植株[37]。稻黃單胞菌(Xan-thomonasoryzaepv.oryzae)是水稻白葉枯病的病原,以gfp為報告基因,利用農桿菌介導法將來自水稻的抗病基因Xa27導入,結果表明Xa27對水稻黃單胞菌具有抗性作用[38]。
展望
在對gfp進行突變改造后,現已獲得發藍光的Bfp、發黃光的yfp和發青色光的cfp,研究人員可根據需要進行選擇[4]。值得一提的是,1999年從珊瑚蟲(discosomagenu)中分離到了與gfp同源的蛋白質dsred,它在紫外線照射下發出紅色熒光,盡管dsred與gfp在氨基酸水平上僅23%的相似度,但三級結構十分相似[12]。與gfp相比,dsred的激發和發射波長較長,發射峰位于培養基、組織培養器材及細胞成分等產生的熒光背景范圍之外,具有較高的信噪比,而且在細胞內熒光轉換效率高,檢測更為容易[39]。隨著研究的進一步深入,一定能開發出更明亮、更穩定、穿透性更強的熒光蛋白,它們將成為生命科學研究的重要工具。
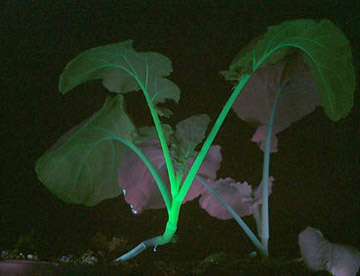
實驗室從青花菜(Brassicaoleraceavar.itali-ca)中克隆到了多個霜霉病(hyaloperonosporaparasitica)抗性相關基因(downymildewresist-ance-relatedgene,dMr),并以gfp為報告基因,對其中的BodMr1進行了遺傳轉化研究。在構建重組載體pBi121-gfp-BodMr1和將其導入農桿菌lBa4404的基礎上,經共養、分化培養、生根培養、熒光檢測和煉苗,成功獲得了17棵轉基因青花菜陽性植株,這些植株在紫外線照射下發出明亮的綠色熒光,抗病性檢測結果證實這些植株對霜霉菌的抗性均有所增加,個別達到高抗水平(待發表)。gfp的引,為快速、有效篩選青花菜轉基因陽性后代及選育抗病材料提供了保障。
 關注我們
關注我們